




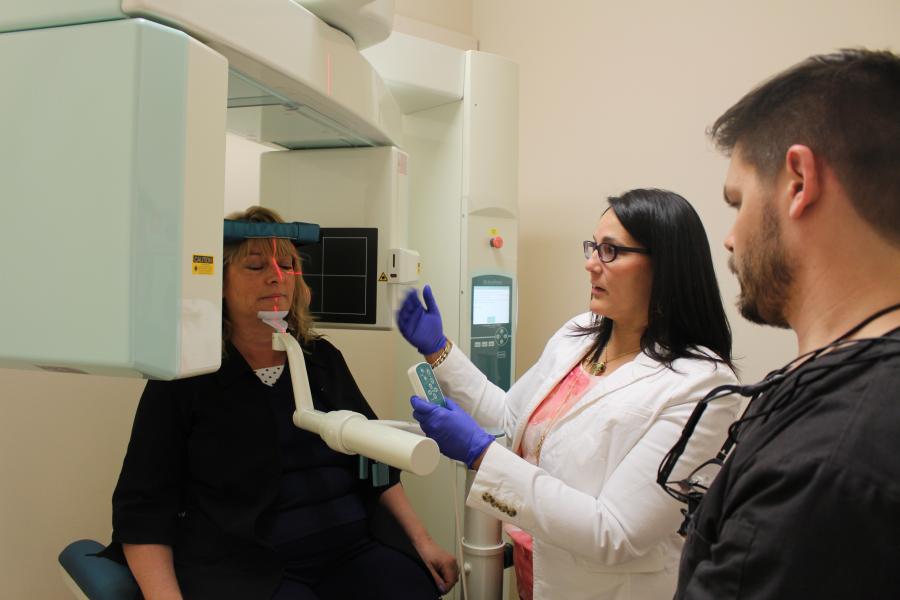

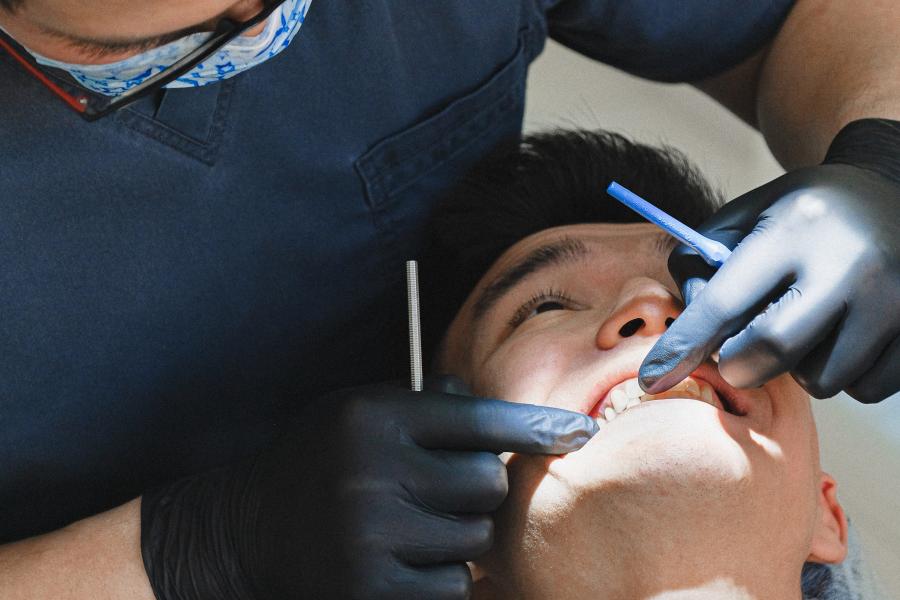
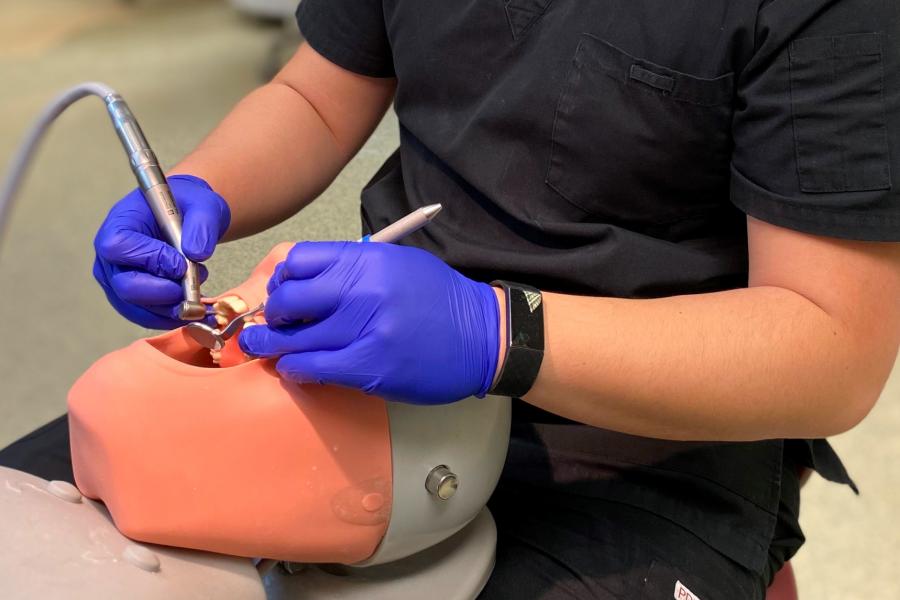
Dental clinic
Our patients have access to general dental services provided by dental students in training. We offer affordable dental care to the public and welcome new patients.
The University of Manitoba campuses are located on original lands of Anishinaabeg, Ininew, Anisininew, Dakota and Dene peoples, and on the National Homeland of the Red River Métis. More
University of Manitoba
Winnipeg, Manitoba Canada, R3T 2N2


Rady Faculty of Health Sciences

Rady Faculty of Health Sciences

Rady Faculty of Health Sciences

Rady Faculty of Health Sciences
Our patients have access to general dental services provided by dental students in training. We offer affordable dental care to the public and welcome new patients.



Apr
2212:00 PM
Online Zoom Session
Apr
2412:00 AM
Apr
2411:00 AM
Online Zoom Session
Dr. Gerald Niznick [DMD/66, LLD (Hon)/02] is recognized as the originator of modern implant dentistry.
Born to immigrant parents, Niznick grew up in Fort Frances, Ontario, before moving to Winnipeg to attend high school, then the University of Manitoba, in the 1960s.
He set up his own dental practice soon after and by the late 80s, Niznick had developed four different implant designs.
His first, the Core-Vent Implant System, in less than a decade became the most widely used dental implant system in the world.
In 2018, Dr. Niznick and Mrs. Reesa Niznick made the largest gift in the College's history.
The Niznicks’ $7.5 million donation reflects the breadth of innovative teaching, research, and community outreach found in the College of Dentistry. Their generous act will enhance clinical training space, fund special initiatives that enrich our community’s health and wellbeing and create reliable endowed funding for the College.
In recognition of this transformative gift, it is a great honour the college bear the name Dr. Gerald Niznick College of Dentistry.

Dr. Gerald Niznick College of Dentistry
D113, 780 Bannatyne Avenue
University of Manitoba, Bannatyne Campus
Winnipeg, MB R3E 0W2 Canada
